
الاتجاهية (علم الأحياء الجزيئي)

الإتجاهية (بالإنجليزية: Directionality) في علم الأحياء الجزيئي وفي الكيمياء الحيوية هي تحديد الاتجاه بين نهايتي سلسلة واحدة لحمض نووي؛ الذي يتم فيه نسخ واصطناع الأحماض النووية والبروتينات.
ترقيم الذرات[عدل]
يتكون الحمض النووي من تكرار لوحدات تسمى نوكليوتيدات وهذه الوحدات تتكون من قاعدة نيتروجينية مرتبطة بسكر خماسي (ريبوز بالنسبة للرنا وريبوز منقوص الأكسجين بالنسبة للدنا) مرتبط بدوره بمجموعة فوسفات.
للتفريق بين ذرات الكربون الموجودة في السكر وفي القاعدة لنيوكليوتيد معين؛ تم الاتفاق على ترقيم ذرات السكر بأرقام تحتوي على فتحة (') لتمييزها عن ذرات القواعد التي يتم ترقيمها بأرقام لا تحتوي فتحة [1] حيث يتم:
- ترقيم ذرات الكربون لحلقة السكر ابتداء من الذرة المرتبطة بذرة الأكسجين من جهة وبالقاعدة الآزوتية من جهة أخرى بالأرقام 1' حتى 5'.
- ترقيم القاعدة الآزوتية من نوع بيريميدين ابتداء من ذرة الآزوت بالاتجاه الأقصر نحو ذرة الآزوت الثانية بالأرقام 1 حتى 6.
- ترقيم القاعدة الآزوتية من نوع بيورين ابتداء بحلقة البيريميدين (السداسية) وانتهاء بحلقة الإيميدازول (الخماسية) بالأرقام 1 حتى 9.
-
 الترقيم القياسي لحلقة الريبوز.
الترقيم القياسي لحلقة الريبوز. -
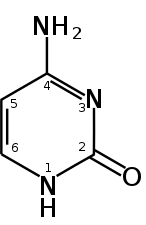 الترقيم القياسي للسايتوسين (بيريميدين).
الترقيم القياسي للسايتوسين (بيريميدين). -
 الترقيم القياسي للأدينين (بيورين).
الترقيم القياسي للأدينين (بيورين). -
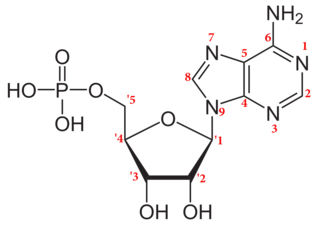 الترقيم القياسي للأدينوسين أحادي الفوسفات (نيوكليوتيد)
الترقيم القياسي للأدينوسين أحادي الفوسفات (نيوكليوتيد)




النهاية 5'[عدل]
النهاية 5' (تنطق النهاية خمسة فتحة) تعني نهاية سلسلة دنا أو رنا تكون فيها ذرة الكربون الخامسة في حلقة الريبوز أو الريبوز منقوص الأكسجين هي الذرة النهائية. يسمح ارتباط مجموعة فوسفات بالنهاية 5' بترابط نيوكليوتيدين وذلك بواسطة الترابط التساهمي بين مجموعة فوسفات النهاية 5' وهيدروكسيل النهاية 3' لنيوكليوتيد آخر لتشكيل رابطة فوسفات ثنائي الأستر. إزالة فوسفات النهاية 5' يمنع ترابط النيوكليوتيدات لمنع تشكل أي حمض نووي غير مرغوب فيه ( مثال الربط الذاتي لناقل بلازميد في التنسيل الجزيئي)، عادة ما يستخدم علماء البيولوجيا الجزيئية الفوسفاتاز لإزالة مجموعة الفوسفات من النهاية 5'.
النهاية 5' لرنا رسول حديث التكوين هي المكان الذي يحدث فيه تقبيع ما بعد الترجمة، وهي عملية أساسية لإنتاج رنا رسول ناضج إذ يزيد التقبيع من استقرار جزيء الرنا الرسول أثناء عملية الترجمة بمنحه مقاومة لعوامل التحلل التي تسببها إنزيمات النوكلياز الخارجية [بحاجة لمصدر]. والتقبيع عبارة عن نوكليوتيدة تمت مثيلتها (ميثيل غوانوسين) مترابطة مع الرنا الرسول برابطة فوسفات ثلاثية 5' - 5' نادرة.
المنطقة الجناحية 5' لجين يقصد بها عادة منطقة دنا لا يتم ترجمتها إلى رنا وتحتوي هذه المنطقة على المحفز الجيني ويمكن أن تحوي كذلك على المعززات ومواقع ارتباط بروتين أخرى.
المنطقة 5' غير المترجمة (5′-UTR) هي منطقة من الجين يتم نسخها إلى رنا رسول وتقع في النهاية 5' لذلك الرنا الرسول، هذه المنطقة من الرنا الرسول قد تتم أو لا تتم ترجمتها إلى بروتين لكن لديها دور في ضبط الترجمة. المنطقة 5' غير المترجمة هي قطعة من الدنا تبدأ من مكان القبعة وتمتد حتى القاعدة التي تسبق AUG رامزة (كودون) بدء الترجمة للتسلسل الرئيسي، يمكن لهذه المنطقة أن تحوي تسلسلات مثل أماكن ارتباط الريبوزوم و تسلسلات كوزاك التي تحدد نجاعة ترجمة الرنا الرسول أو تؤثر في مدى استقراره.
النهاية 3'[عدل]
سُميت النهاية 3' لسلسلة دنا أو رنا كذلك لأنها تنتهي بمجموعة هيدروكسيل الذرة الثالثة لحلقة السكر؛ وتعرف أيضا بنهاية الذيل. هيدروكسيل النهاية 3' ضروري لاصطناع جزيئات أحماض نووية جديد وذلك لأنها ترتبط بفوسفات النهاية 5' لنكليوتيد آخر ما يسمح بإنشاء سلاسل من نيوكليتيدات مرتبطة ببعضها. يستخدم علماء البيولوجيا الجزيئية نيوكليوتيدات لا تحتوي هيدروكسيل 3' (ديوكسي ريبونوكليوتيد) لمقاطعة تضاعف الدنا، تسمى هذه التقنية طريقة إنهاء السلسلة بإنقاص الهيدروكسيل أو طريقة سانغر وتستخدم في معرفة ترتيب تسلسل النيوكليوتيدات في الدنا.
النهاية 3' لرنا رسول ناشئ هي المكان الذي يحدث فيه تذييل ما بعد الترجمة بعديد الأدينيلات والذي يتم فيه ربط سلسلة من 50 إلى 250 جزيء أدينوزين لإنتاج رنا رسول ناضخ، تساعد هذه السلسلة في تحديد مدى بقاء الرنا الرسول في الخلية وهو مايؤثر على كمية البروتين المنتجة منه.
المنطقة الجناحية 3' هي منطقة من الدنا لا يتم نسخها إلى رنا رسول ناضج وهي مجاورة للنهاية 3' للجين المراد نسخه، كان يعتقد في البداية أن هذه المنطقة لا يتم نسخها إطلاقا لكن اكتُشِف بعد ذلك أنه يتم نسخها لرنا ثم تتم إزالتها بسرعة أثناء معالجة النسخة الأولية لتشكيل الرنا الناضج، المنطقة الجناحية 3' تعتوي عادة على تسلسلات لها تأثير على تشكل النهاية 3' للرنا الرسول ويمكن أن تحوي كذلك معزرات ومواقع أخرى ترتبط بها البروتينات.
المنطقة 3' غير المترجمة (3′-UTR) هي منطقة من الدنا يتم نسخها إلى رنا رسول وتقع في النهاية 3' لذلك الرنا ولا تحتوي على تسلسلات رامزات بروتين. يعتبر كل ما هو بين رامزة التوقف وذيل عَدِيْدِ الأدينيلات المنطقة 3' غير المترجمة. يمكن لهذه المنطقة التأثير في نجاعة ترجمة الرنا الرسول أو استقراره كما تحوي تسلسلات مطلوبة لإضافة ذيل عديد الأدينيلات للرنا الرسول، بما في ذلك عديد النوكليوتيد السداسي AAUAAA.
5' إلى 3'[عدل]

لا يمكن تخليق الأحماض النووية في الخلايا الحية سوى في الاتجاهية 5' نحو 3' وذلك لأن البوليميرازات التي تجمِّع مختلف أنواع السلاسل تعتمد على الطاقة الناتجة من كسر روابط نيوكليوسيد ثلاثي الفوسفات لربط نيوكليوسيد أحادي الفوسفات بهيدروكسيل النهاية 3' (-OH) عبر رابطة فوسفات ثنائي الأستر. يُقال عادة عن المواقع النسبية للواحدات على طول سلسلة حمض نووي بما في ذلك الجينات ومختلف مواقع ارتباط البروتينات أنها إما عكس الاتجاه (upstream) أي نحو النهاية 5' أو مع الاتجاه (downstream) أي نحو النهاية 3'.
الإتجاهية لها علاقة بالدلالة لكنها مستقلة عنها، نسخ سلسلة واحدة من الرنا من جزيء دنا مزدوج السلاسل يتطلب اختيار إحدى سلسلتيه لتكون سلسلة القالب التي يتفاعل معها الرنا الناشئ ويتم نسخ تسلسل نيوكليوتيداتها بشكل مكمل. أما السلسلة الأخرى الغير منسوخة فإن تسلسلها مماثل بالضرورة لتسلسل الرنا الناتج، مواقع بدء الترجمة تظهر عادة على كلا سلسلتي جزيء الدنا وتحدد المكان والاتجاه والظروف التي تتم تحتها عملية النسخ. لو كان جزيء الرنا يحمل شيفرة بروتين واحد أو (نادرا) أكثر من واحد فإن ترجمة كل بروتين بواسطة الريبوسوم ستتم في الاتجاهية 5' إلى 3' وستمدد البروتين من نهايته الأمينية حتى نهايته الكربوكسيلية، كمثال: في جين نموذجي كودون البدء (5′-ATG-3′) هو تسلسل دنا يتواجد في السلسلة موجبة الدلالة. يبدأ النسخ عند مكان عكس الاتجاه (بالنسبة للسلسلة موجبة الدلالة) وبينما يمضي عبر تلك المنطقة ينسخ الرامزة 3′-TAC-5′ من سلسلة القالب لإنتاج الرامزة 5′-AUG-3′ داخل الرنا الرسول. يتم فحص الرنا الرسول بواسطة الريبوسوم ابتداء من النهاية 5' أين يُترجم كودون البدء بإدراج الميثيونين (عند حقيقيات النوى) في النهاية الأمينية، بالاتفاق تسلسلات سلسلة واحدة من الدنا أو الرنا مكتوبةٌ في الاتجاهية 5' إلى 3' باستثناء الحالات الخاصة وذلك لتوضيح نمط ترابط القواعد.
انظر أيضا[عدل]
مراجع[عدل]
- ^ DNA - STRUCTURE نسخة محفوظة 16 أكتوبر 2017 على موقع واي باك مشين.
- Harvey Lodish; Arnold Berk; Paul Matsudaira; Chris A. Kaiser (2004). Molecular Cell Biology (5th ed.). New York City: W.H. Freeman and Company. ISBN 0-7167-4366-3.